Were dinosaurs hot-blooded animals?
Viewpoint: Yes, the most recent fossil evidence indicates that at least some dinosaurs were hot-blooded animals.
Viewpoint: No, the long-standing view of dinosaurs as cold-blooded animals is still the most compelling; recent evidence may suggest, however, that some dinosaurs had hybrid metabolisms, or aspects of both hot-and cold-bloodedness.
Naturalists have speculated about the nature of fossils since the time of Aristotle (384-322 B.C. ), but it was not until the seventeenth century that these remarkable objects were generally recognized as the remains of ancient plants and animals. The work of the great French comparative anatomist and geologist Georges Cuvier (1769-1832) established the foundations of pale-ontology, the study of fossilized remains, and stimulated interest in the systematic study of fossils. Richard Owen's (1804-1892) investigations of reptilian fossils led to the discovery that some of these specimens were quite distinct from modern lizards. In 1842 Owen named the new group of large, extinct, reptile-like animals "Dinosauria," meaning "terrible lizard." During the second half of the nineteenth century, many new dinosaur fossils were discovered. Indeed, the 1870s were the beginning of a period known to paleontologists as the "Great Dinosaur Rush." The remains of the great dinosaurs were put on display and drew great public attention.
Since their discovery in the nineteenth century, dinosaurs have been portrayed as gigantic, stupid, sluggish, cold-blooded, lizard-like beasts that flourished in the hothouse world of the Mesozoic period before inevitably passing away and leaving the world to superior warm-blooded mammals. Actually, mammals and dinosaurs coexisted during the Mesozoic era, and, despite modern assumptions about the superiority of warm-blooded mammals, dinosaur predators and herbivores (animals that primarily feed on plant food) dominated the earth as long as they existed. Although the terms "hot-blooded" and "cold-blooded" are often applied to the controversy surrounding the nature of dinosaurs, these terms are not really part of the scientific vocabulary. The proper term for "hot-blooded" is endothermic; this refers to the ability to generate internal heat in order to regulate body temperature. In a cool environment, endothermic animals can maintain an average body temperature that is higher than that of their surroundings. Modern birds and mammals are endotherms. On the other hand, ectothermic animals, such as reptiles, rely on the environment and behavioral adaptations to regulate their body temperature.
A controversial new theory about the nature of dinosaurs was brought to public attention primarily through the work of Robert T. Bakker. Like many pale-ontologists, as a child, Bakker became fascinated by dinosaurs and their place in geological history after seeing dinosaurs in museums and magazine articles. In 1968, just after completing his undergraduate education at Yale, Bakker published "The Superiority of Dinosaurs," his first paper on dinosaur endothermy. Looking at the history of ideas about the evolution of mammals and dinosaurs, Bakker concluded that conventional notions of dinosaur physiology were wrong. Instead of slow-moving reptilian beasts, Bakker envisioned dinosaurs as "fast, agile, energetic creatures" that could run faster than humans. Although critics have labeled one of Bakker's major texts "Dinosaur Hearsay," his articles, lectures, and books, including Dinosaur Heresies and The Great Dinosaur Debate , have been instrumental in stimulating the "Dinosaur Renaissance" proclaimed by Bakker in a 1975 Scientific American article. Since the 1970s, the popular image of dinosaurs has been shifting from cold-blooded reptilian creatures to agile, dynamic, smart, hot-blooded animals. The new image of dinosaurs was reflected in the immensely successful movie Jurassic Park.
John H. Ostrom, Armand de Ricqlés, and other scientists less well known than the flamboyant Robert Bakker have also provided insights into dinosaur ancestry, metabolism, growth characteristics, and behavior. In 1969, Ostrom described a newly discovered theropod dinosaur, Deinonychus antirrhopus , as "an active and very agile predator." Ostrom, who was Bakker's advisor at Yale, was one of the founders of both the theory that birds evolved from dinosaurs and the hypothesis that some, if not all, dinosaurs were warm-blooded. Unlike the continuing controversy about whether dinosaurs were endothermic, the evolutionary relationship between dinosaurs and birds is now generally accepted. The question of which scientist was the first to propose the idea of dinosaur endothermy remains quite controversial.
According to Bakker, evidence for endothermy in dinosaurs appeared in the nineteenth century, but it was ignored because of prevailing assumptions about the reptilian nature of dinosaurs. In 1836 American geologist Edward Hitchcock (1793-1864) studied footprints (subsequently identified as those of dinosaurs) in Massachusetts and suggested that hawk-like creatures that ran made them. Although Richard Owen coined the term "terrible lizards," he seemed to suspect that the dinosaurs might be more warm-blooded than typical reptiles.
Since the debate about hot-blooded dinosaurs began, observers have noted that the case for ectothermy in dinosaurs has generally been taken for granted, rather than demonstrated on the basis of specific evidence. That is, since the ancestors of the dinosaurs were cold-blooded reptiles, the "terrible lizards" were assumed to be ectotherms that functioned quite well because of the warm, mild climate of the Mesozoic era. More recent studies of the climate of the Mesozoic period suggests, however, that the climate was more varied that previously expected and that dinosaurs lived in areas that were fairly cold, such as polar regions. Proponents of the warm-blooded dinosaur hypothesis argue that the discovery of dinosaur fossils at high latitudes supports the theory because ectotherms generally do not live in such cold climates. Nevertheless, modern snakes, lizards, turtles, and crocodiles are quite widely distributed. Moreover, areas in Alaska, Antarctica, and so forth, though cold now, were probably much warmer during the Mesozoic Era. Dinosaur fossils, including hatchlings and juveniles, from the Late Cretaceous (the latest period of the Mesozoic Era) have been found along the North Slope of Alaska, an area that was presumably very cold during the winter season. Thus, the ability of dinosaurs to adapt to cold climates or to seasonal migrations remains a very interesting question.
Some scientists say that the debate about ectothermic and endothermic might be irrelevant. Large dinosaurs could have been "lukewarm," they suggest, rather than "hot" or "cold." Some scientists have proposed an alternative metabolic model for dinosaurs called "Gigantothermy." Because of their immense size, dinosaurs could have been "inertial homeotherms," warming up and cooling down very slowly despite fluctuations in environmental conditions. However, adult dinosaurs ranged in size from as small as 10 pounds to as large as 80 tons, and all dinosaurs would have been quite small as hatchlings and juveniles.
Dinosaur anatomy and posture have been cited as evidence that they were built to move quickly and, therefore, must have had a high metabolic rate. The weight-bearing limbs of dinosaurs gave them a more erect posture than typical reptiles. Critics counter that the evidence does not really tell us how fast dinosaurs moved. Studies of modern ecological patterns suggest that endotherms like mammals and birds usually outcompete and replace ectotherms in any given area. Proponents of dinosaurian endothermy argue that it would have been impossible for mammals and dinosaurs to coexist for about 170 years if dinosaurs had been ectotherms. However, the outcome of competition between modern endotherms and ectotherms has not been established for all possible combinations of species and environments. Thus, the conclusion that endothermic primitive mammals would necessarily have out-competed otherwise highly successful ectothermic dinosaurs should be considered speculative.
Modern ecological studies indicate that the predator/prey ratio that can support mammalian predators is lower than the predator/prey ratios that are needed to sustain ectothermal predators. Based largely on studies of modern mammalian predators and evidence that predator/prey ratios for dinosaurs were apparently quite low, some scientists have argued that dinosaurs must have been endotherms. These studies are interesting, but the calculations depend on many assumptions. In particular, the small numbers of surviving dinosaur fossils mean that sampling errors for both predator and prey species could be very large.
Some advocates of the warm-blooded hypothesis insist that comparison of the structure of dinosaur bones with the bones of mammals, birds, and reptiles provides convincing evidence for dinosaurian endothermy. In cross-section, the bones of dinosaurs more closely resemble those of mammals and birds than reptiles. Because bone morphology within a given species varies considerably depending on age, size, health, and growth conditions, evidence from fossilized bones can be ambiguous. However, sophisticated analysis of dinosaur bones does suggest the rapid growth patterns generally associated with modern endotherms.
Despite increasingly sophisticated studies of dinosaur fossils, paleontologists remain divided about the question of endothermy. More cautious biologists point out that many questions about ectothermy and endothermy in modern animals remain unanswered. Therefore, conclusions made on the basis of fragmentary collections of dinosaur materials should be regarded as highly speculative. During the millions of years of dinosaur evolution, perhaps other alternatives to endothermy and ectothermy appeared. Dinosaurs as a group were very diverse in size, form, habits, and habitat preferences. Presumably, the physiological adaptations of small and large dinosaurs might have been quite different. Dinosaurs dominated the world of terrestrial vertebrates for a period of about 170 million years. Given the long course of dinosaur history, it is also possible that some lines of dinosaurs remained ectotherms, like their reptilian ancestors, while others became endotherms. On the other hand, their physiological adaptations might have been quite different from those of modern reptiles and mammals.
Although several lines of evidence seem to support the hypothesis that dinosaurs were endothermic, critics argue that the evidence is not compelling. Debates about dinosaur physiology are based on a combination of evidence, induction, deduction, and speculation. Dinosaurs cannot be directly compared to any living reptiles, birds, or mammals. Thus, the uncertainties about dinosaur physiology are so great that competing hypotheses are likely to proliferate for many years.
—LOIS N. MAGNER
Viewpoint: Yes, the most recent fossil evidence indicates that at least some dinosaurs were hot-blooded animals.
Because extinct animals—like dead people—tell no tales, scientists determine what their lives and physiology were like by analyzing clues found in fossilized remains and comparing these findings with attributes in living organisms. Over the last few decades, paleontologists have unearthed dinosaur fossils containing well-preserved tissues and organs which indicate that at least some dinosaurs were warm-blooded animals.
In 1968 Robert Bakker, a Yale undergraduate, wrote "The Superiority of Dinosaurs," showing that dinosaurs were "fast, agile, energetic creatures" with an advanced physiology strongly suggesting warm-bloodedness. Bakker showed that 10-ton ceratopsian dinosaurs could outrun a human—an impossible feat for a cold-blooded reptile.
In 1969 Yale professor John H. Ostrom published a paper describing Deinonychus, a theropod dinosaur, as an "active and very agile predator" that had to have a high metabolic rate to sustain active hunting. Bakker's and Ostrom's findings kindled the warm-blooded versus cold-blooded debate.
Metabolism
The terms "cold blooded" or "warm blooded" refer to an animal's average body temperature in relation to its environment. Body temperature is determined by metabolism, the rate at which fuel (food) burns to maintain life.
The technical term for "cold blooded" is ectothermic ( ecto , Greek for "outside"). An ectothermic animal relies on the outside air temperature to regulate its metabolism and body temperature. Ectotherms have low metabolic rates, because food is not used to maintain their internal temperature.
The technical term for "warm blooded" is endothermic ( endo , Greek for "inside"). Endothermic animals (mammals and birds) create heat inside their bodies to maintain body temperature within an optimal range. Endotherms consume lots of food to fuel the inner metabolic engine that maintains their body temperature. Because their bodies burn so much fuel, endotherms have high metabolic rates.
A body's metabolism affects all aspects of its growth and physiology; tissues and organs are formed by and in accordance with metabolism. Dinosaurs' endothermy is established by comparing key characteristics of their physiology and lifestyle with the lifestyle and physiological characteristics common to living endotherms. Each of the following sections describes these comparisons to show that dinosaurs were warm blooded.
Role in Ecosystem
As Top Predators, Dinosaurs Must Have Been Warm Blooded.
All animals metabolize food using the same basic biochemical processes: the same enzymes break down food to yield energy. Metabolic processes are most efficient at an optimal body temperature: for every 10°C drop in body temperature, metabolism becomes twice as slow, producing half as much energy. Thus when it is cold outside, cold-blooded animals are sluggish: their body temperature is too low for efficient metabolism. For this reason, endotherms always outcompete ectotherms.
Evolution teaches that in all cases, long-term competition among organisms in an ecosystem favors endothermic over ectothermic animals. The inability to be active when it is cold or at night is such a disadvantage compared with the endotherm's perpetual internal heat engine that no large cold-blooded predators can dominate mammals.
For more than 140 million years during the Mesozoic, dinosaurs dominated terrestrial ecosystems. The few mammals were tiny rodent-like creatures. Mammals continued to be insignificant, and by the end of the Triassic, dinosaurs occupied nearly every slot in the food chain, from top predator to large herbivore to smaller versions of both. Throughout this period, dinosaurs kept mammals in their insignificant place. At no time could mammals outcompete the dinosaurs. Evolution tells us that this must be because the dinosaurs were endothermic. Had dinosaurs not been endothermic, over these millions of years of evolution the mammals would have outcompeted the dinosaurs and dominated the earth's ecosystems. That did not happen.
Dinosaurs Had Endotherms' Ratio of Hunters to Hunted.
Large warm-blooded predators consume about 10 times more food than cold-blooded predators. The larger the predator, the higher its metabolic rate must be to fuel its huge bulk. Because a large predator needs such huge quantities of food, there are relatively few large predators in any ecosystem. Scientists can calculate precisely the ratio of predators to prey based on the numbers and weight of both.
In today's ecosystems (e.g., African savannah), the ratio of lions (predator) to grazing prey is about 1:100. Studies of prehistoric ectotherms (e.g., Dimetrodon ) show a ratio of 20:100, indicating lower food consumption by cold-blooded animals. Prehistoric mammals (saber-toothed tigers) lived in an ecosystem whose predator-to-prey ratio was about 3 to 5:100. Studies of Tyrannosaurus rex and other Cretaceous dinosaurs also yield a ratio of 3 to 5:100. In all studies, dinosaur predator-to-prey ratios coincided with those of warm-blooded predators and prey.
Distribution: Arctic Dinosaurs
Most dinosaurs are pictured among tropical ferns and cycads. Yet many dinosaur fossils (especially hadrosaurs and theropods) have been found in the Arctic, a region that in the Cretaceous had annual mean temperatures of between 35° and 46°F (2°-8°C). Ectotherms don't thrive in such cold climates. Consequently, almost no ectotherm fossils (e.g., crocodyliform) live in Alaska or other Arctic regions.
Why were dinosaurs able to survive, and thrive, in this cold region when ectotherms could not? The answer must be that the dinosaurs were endothermic.
Some paleontologists suggest that Arctic dinosaurs were ectotherms that migrated seasonally to and from the Arctic. However, Arctic fossil evidence shows the presence of neonates and juveniles, along with adult dinosaurs; this points to year-round habitation. The "migration" theory is questionable, too, because, for all known ectotherms—modern and extinct—a migratory lifestyle is not viable: Their low metabolism does not give ectotherms enough energy for long-term activity.
Bones
Dinosaurs Had Endotherm Bone Structure and Growth.
Bone is a tissue made of a calcium phosphate mineral that is incorporated into a protein called collagen. Collagen occurs in long bundles or fibers.
Dinosaurs had bones that resembled or were identical to endotherm bones in many ways. In ectothermic animals, bone grows slowly. In endotherms bones grow quickly. Fossil dinosaur bones show the rapid growth typical of endotherms.
In ectotherm bones, the collagen bundles are arranged parallel to each other in layers. The resulting bone is dense, with its crystals of calcium phosphate all lined up in the same direction. In endotherms, bones are not dense with neatly aligned fibers but are described as "woven." The collagen is arranged haphazardly, and the calcium phosphate crystals are also oriented randomly. Hundreds of slices of fossilized dinosaur bone, from many different species, show the woven bones typical of endotherms.
Ectotherm bones have pronounced growth rings, because cold-blooded animals grow quickly when it is warm outside and much more slowly when it is cold. Endotherms have bone growth rings, but they are far less pronounced. Dinosaur bones share the poorly defined growth rings found in endotherms.
Ectotherms have relatively few vascular (blood vessel) canals in their bones. Warm-blooded animals have many. Dinosaur bones have the numerous vascular canals common to warm-blooded animals.
Oxygen and Body Temperature.
When bone forms, it incorporates an isotope of oxygen along with calcium phosphate crystals. The amount of oxygen isotope in the bone depends on the body temperature at the time the bone forms.
Analysis of the amount of oxygen isotope in Tyrannosaurus rex bones shows that they were formed while the animal's body temperature was stable within 4°C. Maintaining such a uniform temperature is consistent with the high metabolic rate of warm-blooded animals.
Teeth
In 2001 research described the oxygen isotopes found in 75-million-year-old fossil teeth of crocodiles and theropod dinosaurs. The oxygen isotopes occur in tooth enamel and are not altered during millions of years of fossilization. Fossils of both kinds of animals were obtained from Alaska (cold climate) and Madagascar (hot climate).
The researchers found that the amount of oxygen isotopes in the crocodile teeth varied, depending on the latitude at which the crocodile lived. The high-latitude (cold) crocodiles had relatively little oxygen isotopes in their tooth enamel; isotope levels were higher in those crocodiles from the warmer climate. This variation occurs because in colder climes the reptiles' metabolism is low, so enamel formation is less than in its warm weather cousins.
The dinosaur teeth, however, showed very little variation in the amount of oxygen isotopes, regardless of latitude. Theropods from both sites showed uniform isotope levels, indicating that they maintained a constant body temperature in both cold and warm climates. Thus the dinosaurs must have been endothermic.
Dinosaurs Had Endothermic Organs and Tissues
Brain.
Endotherms tend to have large brains, which function only with a hefty supply of oxygen and fuel (food) and a constant body temperature. Although some dinosaur species did not have large brains, some—particularly the theropods—had brains equivalent to those of similar-sized mammals. This is strong evidence that the theropods were endothermic.
Heart.
In 2001 computed tomography (CT) scans of the innards of 66-million-year-old Thescelosaurus fossils revealed that some soft tissue had been preserved. Imaging of the chest cavity showed a highly evolved heart. The image revealed two adjacent cavities: the lower chambers (ventricles) and a tubelike structure emerging from it (the aorta). Although the thin-walled upper chambers of the heart (the atria) likely collapsed when the dinosaur died, the remaining two chambers and single aorta are identical to the four-chambered heart found in birds and other warm-blooded animals.
Four-chambered hearts evolved in endotherms to fully oxygenate the blood
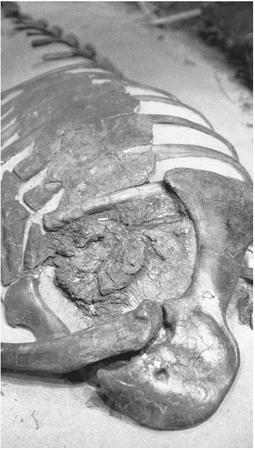
Dinosaur Morphology Supports Endothermy
Bipedalism.
Many dinosaurs—and all theropods—were bipedal: they walked on two legs. Bipedalism requires more energy, and thus a higher metabolic rate, than a more sprawling four-legged posture and gait. Most paleontologists insist that some degree of endothermy is a prerequisite for bipedalism.
Running and Speed.
Modern mammals have an average walking speed far faster than that of modern reptiles. An animal's speed can be calculated by measuring the distance between its footprints. Measurements of fossil footprints show that dinosaurs' speed was equivalent to that of today's mammals. Analysis of Thecodont skeletons, for example, shows that its limbs were built for speed and sustained, rapid running.
To sustain fast and agile running, an animal must have a high metabolic rate. Running is fueled by lungs that efficiently take in large quantities of oxygen and a large heart that pumps this oxygen-rich blood to muscles. Fossils show that most dinosaurs had the wide chest cavity able to accommodate these large organs. Reptiles, and other ectotherms, have much narrower chest cavities incapable of housing large organs. The recently discovered fossilized dinosaur heart (see earlier) confirms this analysis.
Some scientists say that because hadrosaurs and horned dinosaurs do not have wide chest cavities, they were ectotherms. Yet these dinosaurs have air sacs in their vertebrae (back-bone) that are connected to their lungs. Their respiratory system is identical to that found in all modern warm-blooded birds.
Dinosaurs' large hips indicate that they had large leg muscles attached to them—leg muscles that needed lots of oxygenated blood and propelled the dinosaurs at considerable speed. Dinosaur hips resemble those of large-hipped birds and mammals with powerful leg muscles, and they are distinctly unlike the small hips and smaller, weaker reptile leg muscles.
Mating Display.
Few reptiles have evolved displays to attract mates. This makes sense for animals that depend on the outside temperature to jump-start their metabolism. They cannot waste energy on so-called frills.
Yet many dinosaurs evolved to use a great deal of energy on developing displays either to attract mates or repel territorial competitors. Various species of dinosaurs flaunt ornamental crests on their head, huge "sails" on their back, powerful horns for head-butting intruders, and other remarkable—and energy-intensive—"extras." Such morphological "extravagances" are typical of endotherms.
Insulation.
It takes far less energy to heat a well-insulated house than an uninsulated one. Insulation is needed for energy efficiency because the heat is generated inside and must be retained there.
There are absolutely no examples of cold-blooded animals developing body insulation. Insulation is intended to keep heat inside; if the animal obtains heat from outside its body, insulation is counterproductive.
Modern endotherms have various kinds of insulation: blubber, fur, feathers, and down. Some dinosaur fossils reveal the presence of feathers or protofeathers, the forerunner of modern feathers. Recent fossil discoveries in China (e.g., Beipiaosaurus ) indicate that these were indeed feathered, although flightless, dinosaurs. Paleontologists assert that feathers functioned as insulation for these theropod dinosaurs.
Two 120-million-year-old dinosaur fossils found in China in 1998 were unquestionably feathered species. The Caudipteryx had a fan of feathers on its tail and down-like feathers on most of its body. The Protarchaeopteryx was covered with feathers. The find is significant because in both cases the wing feathers were arranged symmetrically, indicating that they were not for flight (birds fly with asymmetrical wing feathers). The researchers therefore concluded that the feathers had to be for insulation.
In April 2001 scientists unearthed a 130-million-year-old fossil dinosaur in China that was covered from head to toe with primitive feathers and down. The bipedal theropod (a dromaeosaur, cousin to velociraptor) yields the strongest evidence yet that dinosaurs are the ancestors of endothermic birds and first developed feathers to insulate their warm-blooded bodies.
Scientists agree nearly universally that dinosaurs are the ancestors of birds. Dinosaurs and birds share many characteristics, including erect posture, similar bone structure, similarly efficient heart and respiratory systems, relatively large brains, the presence of insulating feathers, and care for their young (see later). Birds evolved from dinosaurs in the Jurassic, so it is reasonable to assume that warm-blooded dinosaurs gave rise to warm-blooded birds.
Rate of Dinosaur Evolution Equals That of Endotherms
Reptiles are among the most ancient species on Earth, with some species having remained unchanged for 50 to 60 million years. Because they do not generate internal heat and therefore eat less, ectotherms are far less vulnerable to drastic changes in the environment (e.g., drought) than endotherms. Thus catastrophes put relatively little evolutionary pressure on ectotherms. Their ability to survive adverse conditions accounts for the longevity of ectotherm species.
Warm-blooded animals are highly susceptible to the evolutionary pressures of environmental change. They are also vulnerable to the evolutionary pressure that arises from intense competition. In general, endotherms compete fiercely for resources. They reproduce rapidly and evolve quickly to occupy many different ecological niches. Their ability to diversify, or speciate, quickly to take advantage of newly created niches means that warm-blooded species are relatively short lived. On average, mammals create five or six new genera every 10 million years, compared with reptile species that last from 20 to 50 million years or more. Entire endotherm families survive about 25 million years.
Some dinosaurs—particularly the horned dinosaurs—are known to have generated new species every 5 to 6 million years, just like mammals. Like mammals, these dinosaur families lasted about 25 million years.
Taxonomy tells us that mammals produce more species per genus, and more genera per family, than ectotherms. Scientists know that dinosaurs produced these taxa at a rate similar to that of mammals. For example, dinosaurs averaged 3 to 4 species per genus, and 12 genera per family, the same as mammals. Hadrosaurs and horned dinosaurs created nearly 7 and 6 new genera, respectively, over a period of about 10 million years—again identical to mammals. In contrast, the taxonomic group that includes giant turtles has produced only one new genus in the last 5 million years.
Dinosaurs Had Endotherms' Lifestyle
Hunting.
Most modern reptiles are rather sedentary hunters, simply waiting for prey to blunder by, then lunging and snaring it. The "cold-blooded" lifestyle does not lend itself to active pursuit and hunting.
In contrast, dinosaurs were active hunters. They pursued prey, running relatively long distances on two legs to catch it. Many predatory dinosaurs also had to attack prey and fight long and hard to subdue a large animal with a well-armored body, sharp horns, and other defenses. This level of predatory activity requires endothermy; no ectotherm has the energy for this degree of active hunting.
It is also worth noting that several dinosaur species, particularly Deinonychus , are thought to have hunted at night. Night hunting is almost unknown among cold-blooded animals because nighttime temperatures are generally too low to allow them to be active.
Parental Care.
Some dinosaurs nested in large colonies, where parents incubated the eggs until they hatched. Although this behavior is very much like that of nesting birds, hatching eggs is not, in and of itself, proof that dinosaurs were endotherms. Yet although reptile young hatch from eggs, ectotherms cannot afford to expend their limited energy on feeding their off-spring once they have hatched. Once born, reptile young are on their own.
Studies of Maiasaura nesting areas have yielded evidence that after hatching, the young remained in the nest for a significant period of time. The fossil evidence includes eggshells repeatedly trampled by the young, and fossils of juveniles of different ages found in the nest. From this evidence, scientists stipulate that the dinosaur parents must have gathered food and brought it back to feed their growing young in the nest—behavior common only to endotherms.
Conclusion
Overwhelming evidence indicates that at least some dinosaur species were endothermic. There can be no doubt, based on the data and their clearly endothermic characteristics, that some dinosaur species were warm blooded, with some form of endothermic metabolism.
—NATALIE GOLDSTEIN
Viewpoint: No, the long-standing view of dinosaurs as cold-blooded animals is still the most compelling; recent evidence may suggest, however, that some dinosaurs had hybrid metabolisms, or aspects of both hot-and cold-bloodedness.
Despite recent media depictions of dinosaurs as warm-blooded animals, evidence strongly favors the hypothesis that the long extinct creatures of the Mesozoic era were cold-blooded organisms. Although the decades-old notion that dinosaurs were cold blooded is still sound, many scientists are now considering the possibility that dinosaurs may, in fact, have had a so-called hybrid metabolism that would have given them the best of both worlds. They would have had the benefits of a warm-blooded animal, complete with on-demand explosive energy spurts, while enjoying the much lower energy requirements of their cold-blooded lifestyle the rest of the time. Hybrid or not, the evidence for cold-bloodedness is persuasive.
Cold-bloodedness (the scientific terms are ectothermy or poikilothermy ) means that an animal's internal or body temperature fluctuates with the outside temperature. For example, a dragonfly, grasshopper, or other insect is ectothermic, so it is sluggish on a cool summer morning but becomes increasingly active as the day warms. The temperature rises, the insect warms, the chemical reactions that control its metabolism speed up, and the insect can move more quickly. The same process happens in amphibians (like frogs and salamanders) and reptiles, all of which are ectotherms.
Historically, the scientific community has placed dinosaurs in the cold-blooded category because of their close phylogenetic relationship with reptiles, which include turtles, lizards, snakes, crocodiles, and alligators. Based on overall bone structure and other clues from the fossil record, scientists have concluded that dinosaurs evolved from reptiles. Because dinosaurs branched off the reptilian evolutionary tree, it naturally followed that dinosaurs were cold blooded like their ancestors, and that hypothesis has yet to face any conclusive evidence to the contrary.
The greatest hurdle in the debate of warm-bloodedness (also known as endothermy or homeothermy ) versus cold-bloodedness is the lack of information. The fossil record, although growing, is far from complete. Paleontologists frequently find only a few broken pieces of dinosaur bones and then have the daunting task of making inferences about physiology, behavior, and social structure from these tiny remnants. On those rare occasions when scientists unearth an entire skeleton, they often struggle to answer definitively even the basic questions: Was it male or female? What did it look like? How did it live? In light of these most fundamental uncertainties, cold-blooded proponents typically see the warm-blooded argument as little more than a collection of ambiguous findings and ungrounded conjecture.
Hearts and Heat Loss
For example, warm-blooded supporters frequently tout the highly publicized discovery of a fossilized dinosaur heart in 1993 as proof of their claim. Fossil collector Michael Hammer of Augustana College in Illinois made the find in South Dakota among the bones of a 660-pound, 13-foot-long, 7-foot-tall Thescelosaurus, a plant-eating beast that roamed the earth 66 million years ago.
Soon after he announced his discovery, scientists began questioning whether the grapefruit-sized clump of red rock was actually a heart at all: The chest (thoracic) cavity contained none of the other preserved soft tissues that, they said, should have also been present; and some of the purported heart was actually outside the cavity rather than inside where it should be.
Even if the structure was cardiovascular tissue, the cold-blooded camp rejected the Hammer group's conclusion that it originated in an endothermic animal. Using computed tomography (CT) scans to see inside the find, Hammer's group suggested the heart's structure matched that of warm-blooded animals, like mammals and birds: it had two lower chambers, or ventricles, that separated the oxygenated, inhaled air from the exhaled air, but it had only one aorta, the main blood vessel leading out of the heart.
Reptiles, in contrast, have two aortae, and most have a three-chambered heart with two upper chambers (atria), but just one ventricle. Of Hammer's finding, cold-blooded proponents noted that the fossil was incomplete, lacking the atria as well as carotid and pulmonary vessels. Likewise, they asserted that a second aorta may well have been present, but not preserved.
In addition, the presence of a second ventricle does not exclude ectothermy, because some reptiles have four-chambered hearts. The modern cold-blooded monitor lizard, for example, exhibits two-phased pumping in its ventricle, so its heart functions as a four-chambered organ. Crocodilians (crocodiles, alligators, and their allies) have a completely divided ventricle and thus a four-chambered heart.
With this reasoning, cold-blooded supporters contended that even if the Hammer group's description of the heart is correct, it still comes nowhere near proving that the dinosaur was warm blooded, only that it had a two-ventricle, one-aorta heart.
Some warm-blooded supporters also use the allegation that cold-blooded animals could not have survived in particularly cold climates, because they could not extract enough heat from the environment to support them. However, many present-day reptiles survive quite well in extreme temperatures.
A 1990 study on leatherback turtles showed how dinosaurs could cope with the temperature extremes. For this study, the scientists investigated how the large turtles regulate their body temperatures on their long migrations from the hot tropics to the downright cold North Atlantic waters. They found that the turtles use their size—some tip the scales at more than 1,500 pounds—and insulating tissue to maintain a fairly constant internal temperature, even in highly variable weather conditions.
The researchers, whose work appeared in Nature, extrapolated from the turtle's present-day abilities and concluded that dinosaurs could have had a similar capacity to survive in wide-ranging climates. According to Drexel University's James Spotila, one of the authors of the research, "Because of their large body size, dinosaurs were able to use their bulk as insulation and as a countercurrent heat exchanger." He added, "At a certain bulk level, warm-blooded and cold-blooded aren't that much different."
Clues in a Dinosaur's Breath
Work published in 1997 and 1999 provided a glimpse of dinosaur metabolism and helped confirm the cold-blooded hypothesis. In these studies, scientists examined fossils from various dinosaurs.
In the 1997 study, the researchers studied the nasal bones of three different species: the ostrich-like Ornithomimus, a duck-billed Hypacrosaurus, and the Tyrannosaurus relative called a Nanotyrannus.
Using scanning equipment, they found that the dinosaurs lacked the coiled cartilaginous or bony structures, called respiratory turbinates, that are found in nearly all endotherms, but not in ectotherms. Actually, the scientists looked for the bony ridges that support the turbinates rather than the coiled structures themselves, because turbinates are delicate and rarely survive fossilization. They readily found the telltale ridges in fossilized mammals, but not in dinosaurs. The dinosaurs also exhibited narrow nasal passages similar to those in present-day reptiles. Endotherms, however, require large passages to house the turbinates and also to admit the greater amount of oxygen required by a warm-blooded animal.
Lead researcher John Ruben, a professor of zoology at Oregon State University at Corvallis, said, "The nasal structures we saw in dinosaur fossils simply would not have been able to accommodate the higher lung ventilation rates that dinosaurs would have had if they were warm-blooded."
In the 1999 research, Ruben's group added fuel to the ectothermy supposition following an analysis of a fossil of a baby Scipionyx samniticus, a small, velociraptor-like, meat-eating dinosaur. Using ultraviolet light to see the fossil in detail, they found and examined internal organs that had been preserved along with the fossil. Most importantly, they discovered that the lungs and heart were located in one part of the body cavity and the liver in another part. A diaphragm separated the two cavities. This division is significant, because it only occurs in animals that use a diaphragm to improve the lungs' performance.
By studying the liver's structure and the muscles surrounding it, the researchers surmised that dinosaur respiration was similar to that of the crocodilians living on Earth today. In crocodilians, abdominal muscles drive the liver up into the lungs to force air into and out of the lungs, much like a bellows draws in air, then blows it out to stoke a fire. Likewise, the Scipionyx had large muscles that turned the liver into a piston-like device for improving respiration when the animal demanded a higher metabolism. Such a system would allow dinosaurs the bursts of high metabolism needed to chase prey, escape from a predator, or engage in other high-energy activities.
Team member and paleobiologist Nicholas Geist remarked, "These theropod dinosaurs were fast, dangerous animals, certainly not slow or sluggish. They could conserve energy much of the time and then go like hell whenever they wanted to." Besides the liver-aided exhalation, the research team also noted that the fossil contained simple lungs that are much more similar in appearance to modern crocodilians than to warm-blooded animals.
The Hybrid Notion
Although many scientists are convinced dinosaurs were not warm blooded, they are becoming increasingly intrigued by another notion: the animals may actually fall somewhere between warm and cold blooded. This hybrid metabolism would allow dinosaurs to be as active as endotherms when necessary, but revert to the much less energy-demanding lifestyle of an ectotherm when at rest. In support of this viewpoint, the scientists point to fossil findings and research showing that although dinosaurs are very close to reptiles, they are not identical.
A 1998 study of growth rates is a case in point. Kristina Curry of the State University of New York at Stony Brook analyzed the growth

She based her conclusion about age, however, on the number of annual growth rings present in cross sections of the animal's bone, and the growth rings themselves are indicative of reptiles, not mammals. Ectotherm bones display their age by these visible annual lines of arrested growth, called LAGs, because colder seasonal temperature slows their metabolism and, therefore, their growth every year. Modern endothermic bone only acquires LAGS if the animal experiences some type of stress, such as starvation. In summary, the simple presence of the growth rings implies the animals were ectothermic, and the swiftness of development only indicates that the sauropods grew to maturity much more quickly than formerly assumed and at a speed similar to endotherms.
Although the media hype of warm-bloodedness has been in full force recently, the bank of scientific evidence has quietly tilted in favor of dinosaurs having a cold-blooded or perhaps a hybrid metabolism. Just because cold-blooded supporters endorse the traditional view of dinosaur metabolism, it does not mean they are not open to new ideas.
Most, in fact, no longer view dinosaurs as not particularly bright reptiles that slowly plodded through the terrain of the Mesozoic era. Instead, they believe dinosaurs were smarter (although not on the level of the overly intelligent velociraptors in Jurassic Park ), more active and something other than just big lizards. With their combination of ectothermic metabolism and wide activity range, said paleontologist James Farlow at Indiana University-Purdue University, "It's not surprising that they ruled Earth for over 100 million years."
—LESLIE MERTZ
Further Reading
Associated Press and United Press International."Huge Turtles May Show How Dinosaurs Endured Hot, Cold." Detroit Free Press (April 26, 1990): 2A.
Bakker, Robert. Dinosaur Heresies. New York:Kensington, 1986.
——. The Great Dinosaur Debate. New York:Citadel Press, 2001.
"Dinosauria." University of California at Berkeley, Museum of Paleontology. <www.ucmp.berkeley.edu/diapsids/dinosaur.html> .
"The Evidence for Ectothermy in Dinosaurs." <www.ucmp.berkeley.edu/diapsids/ectothermy.html> .
Gibbs, Ann. "Lung Fossils Suggest DinosBreathed in Cold Blood." Science 278 (November 1997): 1229-30.
May, Julian. The Warm-Blooded Dinosaurs . NewYork: Holiday House, 1978.
Morell, Virginia. "A Cold, Hard Look atDinosaurs." Discover 17 (December 1996): 98-102.
"New Insights on Dinosaur Metabolism." USA Today (June 1999): 8-9.
Ostrom, J.H. "Terrestrial Vertebrates as Indicators of Mesozoic Climate." Proceedings of the North American Paleontological Convention (1969): 34-76.
——. The Evidence for Endothermy inDinosaurs. AAAS Symposium 28 (1980): 15-52.
Ross-Flanigan, Nancy. "The Diversity ofDinosaurs: The More Scientists Find, the More They Find Differences." Detroit Free Press (June 1, 1993): 1D.
Ruben, John. "Pulmonary Function and Metabolic Physiology of Theropod Dinosaurs." Science 283 (January 1999): 514-6.
Thomas, Roger D. Cold Look at Warm-Blooded Dinosaurs. Boulder, CO: Westview Press (AAAS), 1980.
Weed, William Speed. "What Did DinosaursReally Look Like—and Will We Ever Know?" Discover 21, no. 9 (September 2000): 74-81.
Wuethrich, Bernice. "Stunning Fossil ShowsBreath of a Dinosaur." Science 283 (January 1999): 468.
KEY TERMS
ECTOTHERM:
"Cold-blooded" animal; an animal that relies on the outside air temperature to regulate its body temperature; ectotherms (animals that are ectothermic) have a low metabolic rate.
ECTOTHERMY (POIKILOTHERMY):
Also called cold-bloodedness; metabolic state by which an animal's internal or body temperature fluctuates depending on the temperature of the surrounding environment.
ENDOTHERM:
"Warm-blooded" animal; an animal whose high metabolic rate maintains a fairly constant internal body temperature.
ENDOTHERMY (HOMEOTHERMY):
Also called warm-bloodedness; metabolic state by which an animal's internal or body temperature remains relatively stable regardless of the temperature of the surrounding environment.
ISOTOPE:
Variation on the most common form of the atom of any element; most isotopes occur because of a different number of neutrons in the atomic nucleus.
LINES OF ARRESTED GROWTH (LAGS):
Annual growth rings seen in the bones of ectotherms but not endotherms.
METABOLISM:
Rate at which food is "burned" as fuel inside the body.
MORPHOLOGY:
The form, or shape, of a body and its parts.
PALEONTOLOGIST:
Scientist who studies prehistoric forms of life through its remains in plant and animal fossils; a scientist in the field of paleontology.
PREDATOR:
Any animal that hunts and eats other animals.
PREY:
Any animal that is hunted and eaten by other animals.
RESPIRATORY TURBINATES:
Coiled cartilaginous or bony nasal structures found in endotherms but not ectotherms.
SPECIATE:
To create a new species; the process in which new species are created through evolution (speciation).
TAXONOMY:
The scientific system of classification, particularly of living things, that are grouped according to common characteristics; taxa (sing. taxon): a particular classification of living things; taxa mentioned in the text include from the broadest to the narrowest classification of organisms: family, genera (sing., genus), and species.
THEROPOD:
Any of a group (Theropoda) of bipedal, carnivorous dinosaurs.
GIGANTOTHERMY
Some scientists suggest that when animals become extremely large, entirely new processes come into play to maintain an internal body temperature. The argument for gigantothermy can be explained through a simple analogy.
Imagine a bathtub filled to the top with hot water. Then imagine a wide shallow soup bowl also brimming with hot water. In the case of the bathtub, only the water on the surface is exposed to the air that cools it. The vast volume of water in the tub is not exposed to the air, so does not cool as readily. In the case of the soup bowl, the water surface exposed to the cooling air is relatively large compared to the volume of water in the bowl. Thus the relatively small volume of water beneath the surface will cool rather readily. The key difference in these two examples is the surface-to-volume ratio. The lower the surface-to-volume ratio (the case of the bathtub), the more inner heat is retained.
Obviously, dinosaurs had a low surface-to-volume ratio. Their skin covered an enormous gut that was far more voluminous than the skin surface. Thus scientists have argued that the dinosaurs would lose little internal heat to the outside. The sheer bulk of the dinosaur innards was sufficient to maintain a fairly stable internal temperature.
But several problems arise with this argument. First, gigantothermy is generally able to maintain an internal temperature within 6° to 8°C. Fossil studies have shown that some dinosaurs maintained their internal temperature within 4°C. This range of temperature stability, although larger than that seen in modern mammals, is indicative of some degree of endothermy.
Second, as discussed in the text, the dinosaurs outcompeted the mammals with whom they lived for many millions of years. Had the dinosaurs been gigantothermic, evolution tells us that the warm-blooded mammals would have outcompeted them. They did not.
Third, dinosaurs' ancestors were relatively small animals for whom gigantothermy was impossible. Over time, evolution allowed these small ancestors to dominate the small mammals that also lived then. For the small dinosaur ancestors to have out-competed the mammals, these ancestors must have been endothermic—and endothermy served them well in the competition for dominance. Why then, would the descendants of small endothermic animals abandon dominance-conferring endothermy for gigantothermy? In evolutionary terms, there would be no reason for the dinosaur descendants to give up a winning strategy.
Finally, we must consider the fact that some dinosaurs lived in relatively cold climates. Although adult dinosaurs might have been able to tolerate cold conditions through gigantothermy, there is no way their young could have done so. Dinosaur hatchlings were small—too small for gigantothermy. Had they not been endothermic, the young would have died and no dinosaurs would have been found in these cold regions.
—Natalie Goldstein
Nate's comment or question about a dinosaur in an icey lake prompted this search. I will also forward one, more scientifically written. Both very informative,this one is easier to follow, I must say. Lisa
Thanks!